Общий стереохимический генетический код - путь к биотехнологии и универсальной медицине XXI века уже сегодня
Л. Б. Меклер, Р. Г. Идлис
Раздел 6
САМОСБОРКА ПРОСТЕЙШЕЙ БИОЛОГИЧЕСКОЙ МИКРОМАШИНЫ: УЧИМСЯ ЯЗЫКУ БИОЛОГИИ
Подавленные утратой необходимости их экспериментального искусства, - результата появления на рынке широкого ассортимента автоматизированных наборов инструментария и реактивов для молекулярно-биологических исследований, - утратой самой цели этих исследований, - результата накопления в общедоступных электронных банках данных все большего числа нуклеотидных последовательностей-генов, - молекулярные биологи-практики восклицают: "Молекулярная биология погибла - Да здравствует молекулярная биология!"...молекулярная биология новой парадигмы, цель которой - разработка теории, проверяемой экспериментом".
К сдвигу парадигмы биологии.
Уолтер Гилберт, лауреат
Нобелевской премии
(Nature. 1991. V. 349. Р. 99)
Ниже на примере тахиплезина, полипептида, защищающего крабов от бактерий, мы продемонстрируем, как, исходя из разработанной нами теории, можно за считанные минуты проявить смысл генетического текста, кодирующего и строение, и программу работы этой маленькой биологической микромашины, т. е. вручную, без всякого компьютера, построить и А-А-модель, и П-К-модель этой трехмерной молекулы.
Примечательно, что попыткам понять принципы проектирования Природой таких биологических микромашин и принципы их работы сейчас посвящают целые симпозиумы
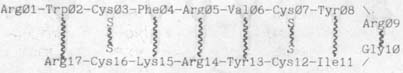
35.Тахиплезин - полипептид, состоящий всего из 17 аминокислотных остатков, четыре из которых - цистеины
36.Хорошо известно, что цистеины, взаимодействуя друг с другом, образуют ковалентные связи, так называемые S-S-мостики. Именно эти S-S-мостики и придают данным трехмерным молекулам ту степень жесткости, которая необходима для выполнения ими их физиологических функций. Например, в полипептидной цепи выделенного из сои Боуманом и Бирком ингибитора трипсина и химотрипсина из числа ее 71 аминокислотного остатка 14 - цистеины. Именно семь S-S-мостиков, образуемых этими цистеинами, и ответственны за способность этого ингибитора, заполняя полости активных центров этих ферментов подобно пробке, лишать их подвижности - свойства, совершенно необходимого для функционирования любого фермента. Нетрудно подсчитать, что чисто формально 14 цистеинов могут образовать 13x11x9x7x5x3x1=135135 вариантов сочетаний семи S-S-мостиков этого ингибитора. В действительности, однако, реализуется только один из этих 135135 вариантов, и всегда безошибочно!
Как же Природа решает эту задачу? Чтобы ответить на этот вопрос, мы построили, согласно данной теории, - вручную модель трехмерной молекулы этого ингибитора и, чтобы проверить, насколько она верна, сравнили ее с моделью этого полипептида, построенной по результатам ЯМР-анализа его концентрированного раствора
37.Результаты сравнения показали, что эти модели адекватны друг другу не только по положению S-S-мостиков, но и топологически - по общему ходу в пространстве полипептидной цепи. Вероятность случайного получения этого результата - даже только в отношении предсказания положения в этой макромолекуле ее S-S-мостиков ~10-6.Именно имея в виду небеспредельные рамки данной статьи, далее ограничимся построением моделей трехмерной молекулы тахиплезина (
рис. 4). Формально при формировании трехмерной А-А-молекулы тахиплесина могут реализоваться три варианта ее двух S-S-связей. Сегодня молекулярная биология, исходя из аминокислотной последовательности даже такого маленького полипептида, ничего не может сказать ни о его трехмерной структуре вообще, ни о положении его S-S-связей в частности. Ибо огромное число степеней свободы этой полипептидной цепи исключает возможность рассчитать ее конформацию согласно законам физики или химии, например, исходя из величин энергии взаимодействий ее атомов.Согласно теории, которую мы разработали, трехмерная структура любого полипептида определяется биологически - совокупностью А-А-связей, образующихся между его аминокислотными остатками. Обращаясь к графу А-А-кода (см.
рис. 2 и табл. 2), видим, что аминокислотные остатки тахиплезина формально могут образовать друг с другом только пять следующих А-А-связей: Tyr8-Val6, Tyr13-Ile11, I1е11-Туг8, Туг13-Val6 и Lys15-Phe4 (они показаны в табл. 2 скобками). Обращаем внимание читателей, что две из этих А-А-связей, Туг8-Val6 и Туг13-I1е11 (показаны скобками, стоящими слева от соответствующих аминокислотных остатков), не могут реализоваться, ибо А-А-связи между аминокислотными остатками, порядковые номера которых отличаются друг от друга на 1 или на 2, стерически невозможны. Чтобы убедиться в этом, достаточно попытаться построить их модели из моделей атомов, разработанных Л. Полингом, Р. Кори и У. Л. Колтуном (так называемых full space filling CPK atomic models) (СРК-моделей)38.Реализация оставшихся трех А-А-связей, показанных в табл. 2 квадратными скобками, стоящими справа от соответствующих аминокислотных остатков, стерически возможна и приводит к тому, что полипептидная цепь тахиплезина складывается пополам (см.
рис. 4б и схему). В результате Cys12 оказывается напротив, т. е. в суперпозиции к Cys7, а Cys16 - напротив, т. е. в суперпозиции к Cys3. Согласно данной теории, это означает, что S-S-связи этой трехмерной молекулы должны образоваться именно между Cys12 и Cys7 и между Cys16 и Cys3. Результаты химического анализа тахиплезина, опубликованные в литературе, демонстрируют, что в его трехмерной молекуле действительно содержатся S-S-связи Cys12-Cys7 и Cys16-Cys3 (см. табл. 2).Чтобы убедиться в том, что модель трехмерной молекулы тахиплезина, построенная согласно данной теории из бумажной ленты - имитатора полипептидной цепи тахиплезина, тем не менее построена в полном соответствии с законами стереохимии мы построили данную модель также из СРК-моделей ее аминокислотных остатков, имея в виду, что в этом случае все соответствующие законы стереохимии будут выполняться автоматически (см.
рис. 4г).Обращаем внимание читателей, что СРК-модели этих биополимеров мы строим не из атомных моделей соответствующих полипептидных цепей, а из готовых атомных моделей тех А-А-связей, которые, согласно данной теории, образуются по ходу формирования трехмерной молекулы данного биополимера. Такая последовательность построения их моделей и возможна, и необходима, и достаточна. Возможна - потому, что стереохимическая архитектура А-А-связей столь же характерна и индивидуальна, сколь и стереохимическая архитектура ковалентных связей, например, молекул Н
2О,H3PO4, KMnO4, CH4, NН3 и т. д. Необходима потому, что при таком построении лишний раз проверяется правильность построения атомных моделей А-А-связей (ибо если вместо данной А-А-связи пытаются в данную полипептидную цепь встроить, например, даже ее минимальную альтернативу, скажем, вместо А-А-связи Туг-Ilе, А-А-связь Туг-Leu, то такую альтернативную пару, хотя и построенную по тем же законам стереохимии и термодинамики, тем не менее, судя по нашему опыту, как правило, состыковать с соответствующими аминокислотными остатками не удается). Достаточна - потому, что служит доказательством верности модели, построенной из бумажной ленты.В данном случае мы взяли готовые СРК-модели А-А-связей Ilе-Туг, Туг-Val и Lys-Phe и состыковали аминокислотные остатки (компоненты этих А-А-связей) с остальными соответствующими аминокислотными остатками полипептидной цепи тахиплезина, воссоединив все ее пептидные связи. В результате мы убедились, что в А-А-модели трехмерной молекулы тахиплезина, построенной из СРК-моделей ее аминокислотных остатков, именно Cys12 иCys7, а также Cys16 и Cys3 находятся в суперпозиции друг к другу, т. е. именно в том положении, при котором S-S-связи могут образоваться только между данными Cys, что и требовалось доказать: именно последовательное образование, согласно данной теории, соответствующей совокупности А-А-связей приводит в суперпозицию именно те аминокислотные остатки Cys, между которыми S-S-связи выявлены в реальной молекуле тахиплезина.
Видим далее, что все аминокислотные остатки этой сложенной пополам полипептидной цепи, находящиеся в суперпозиции друг к другу, стереокомплементарны друг другу согласно П-К-коду. Напоминаем, что, согласно данной теории, формирование "твердых", "нативных" трехмерных молекул полипептидов и белков детерминировано П-К-кодом ОСГК. В данном случае это означает, что непрерывная последовательность из этих восьми П-К-связей представляет собой не что иное, как защитную рубашку совокупности -NH...OC- водородных связей, образовавшихся между аминокислотными остатками, стоящими в суперпозиции друг к другу. Следовательно, трехмерная молекула тахиплезина, находящаяся в П-К-конформации, представляет собой, по сути дела, антипараллельную b-структуру, образованную сегментами Тгр2-...-Аrg9 и Аrg17-...-Gly10 (см. рис. 4 в).
Известно, что тахиплезины представляют собой семейство гомологов, отличающихся друг от друга на 1-2 аминокислотных остатка (Аrg1 может быть заменен на Lys1, a Lys15 - на Аrg15). Видим, что, хотя эти замены могут привести к исчезновению А-А- и П-К-связей Lys15-Phe4 в случае, когда Lys15 заменен на Аrg15 (что, согласно данной теории, у молекулы [Аrg15]-тахиплезина должно привести к замедлению образования ее S-S-мостика Cys16-Cys3), возникновения в "защитной рубашке" данной b-структуры этой единственной "дырки" недостаточно, чтобы эта b-структура распалась. Следовательно, антипараллельная b-структура в рамке, образованной данными восемью или семью П-К-связями, должна содержаться в трехмерных молекулах всех этих гомологов семейства тахиплезина - таково предсказание данной теории.
Сравнивая эту модель трехмерной молекулы тахиплезина с моделью одного из его гомологов ([Lys1, Агд17]-тахиплезина), построенной по результатам ЯМР-анализа его концентрированного раствора39, убеждаемся, что эта ЯМР-модель тоже представляет собой антипараллельную b-структуру,образованную сегментами Cys3-...-Туг8 иCys16-...-I1е11, т. е. b-структуру в границах данной защитной рубашки, что и требовалось доказать.
Итак, мы видим, что при построении согласно данной теории моделей даже столь маленького полипептида, как тахиплезин, удается однозначно не только выявить всю последовательность событий, ведущую к формированию сначала "жидких", а затем и "твердых" конформеров этих биополимеров, но и увидеть невооруженным глазом их взаимное превращение друг в друга - основу всей биомеханики. Более того, столь же наглядно удается убедиться и в том, что образование S-S-связей этих биополимеров - действительно прямое следствие формирования предшествующей совокупности соответствующих А-А-связей этой молекулы.
И удалось достигнуть этих целей столь просто только потому, что, ведя эту работу, мы задавали вопросы Природе не на языке физики, не на языке химии, а на языке биологии.
Конечно, построение согласно данной теории моделей других, не только больших, но и столь же малых полипептидов, часто более трудная задача, выполнимая только в том случае, когда она решается согласно соответствующему алгоритму, центральная задача которого - отбор среди альтернативных вариантов А-А-связей энергетически наиболее выгодных. И, стремясь вывести на первый план построения этих моделей самую суть этого процесса в чистом виде, не осложненную отбором вариантов А-А-связей, мы поэтому и избрали для его иллюстрации построение моделей трехмерных молекул именно этого тахиплезина.
Что касается данного алгоритма, то иллюстрация его работы - содержание очередной статьи, посвященной построению трехмерной молекулы другого ингибитора, так называемого
m-конотоксина. Мы, хотя бы максимально кратко, говорим об этом в данной статье только потому, что построение согласно данной теории моделей трехмерной молекулы m-конотоксина снимает завесу тайны с, по-видимому, очень важного биологического явления, экспериментально неожиданно обнаруженного Т. Крейтоном еще 20 лет назад при попытке выявить механизм обратимой ренатурации панкреатического ингибитора трипсина крупного рогатого скота, а тем самым и механизм формирования трехмерных молекул белков вообще40. Имеется в виду тот факт, что походу этого процесса сначала образуются совсем не те три S-S-связи этой трехмерной молекулы, которые существуют в ней, судя по результатам РСА кристалла этого ингибитора. С тех пор этот процесс интенсивно изучается по обе стороны Атлантики, но без особого успеха: ни причин рекомбинации этих Cys-S-S-Cys-связей, ни последовательности их рекомбинации в результате этих экспериментов до сих пор однозначно установить не удалось. Ибо вопросы этими экспериментаторами задаются Природе на языке химии, а не на языке биологии. Pollard Т. D. // Nature. 1992. V. 355. Р. 17-18; Proteins as machines // Indiana University, 27-30 October, 1991; Hoffman M. Motor Molecules on the Move ("The cell is not just filled with randomly moving parts but is well equipped with special proteins that power internal movements") // Science. 1992. V. 256. P. 1758-1760; Alberts В., Miake-Lye R. Uncrambling thePuzile of Biological Machines: The Importance of the Details // Cell. 1992. V. 68. P. 415-420. Nakamura T. et al. // J. Biol. Chem. 1988. V. 263.P. 16709-16713; Miyata T. et al. //J. Biochem. 1989.V. 106. Р. 663-668. Werner M. H., Wemmer D. Е. // Biochemistry.1992. V. 31. Р. 999-1000. Ко11un W. L. Biopolymers. 1965. V. 3. Р. 665-679. Кawanо К. et al. // J. Biol. Chem. 1990. V. 265.Р. 15365-15367. Creighton T. Е. Understanding protein folding pathways and mechanisms // PROTEIN FOLDING. Deciphering the Second Half of the Genetic Code / Ed. by L. M. Gierasch and J. King // American Association for the Advancement of Science. 1990. P. 157-170; Creighton T. Е. Up the kinetic pathway // Nature.1992. V. 356. P. 194-195.Титульный лист | Физико-химическая биология | Меклер
{kind=link}